Célula del fotorreceptor
| Célula del fotorreceptor | |
|---|---|
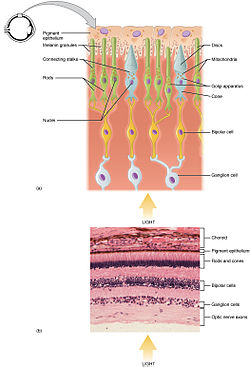
Partes funcionales de la barras de y conos de, que son dos de los tres tipos de células fotosensibles en la retina
|
|
| Identificadores de | |
| NeuroLex ID | Célula del fotorreceptor |
| Terminología anatómica | |
A célula del fotorreceptor es un tipo especializado de neurona encontrado en el retina es capaz de fototransducción. La gran importancia biológica de los fotorreceptores es que convierten la luz (visible radiación electromagnética) en señales que pueden estimular los procesos biológicos. Para ser más específicos, proteínas del fotorreceptor en la célula de absorber fotones, provocando un cambio en la célula potencial de membrana.
Son las dos células del fotorreceptor clásico barras de y conos de, cada uno aportando información utilizada por el sistema visual para formar una representación del mundo visual, vista. Las barras son más estrechos que los conos y distribuidos diferentemente a través de la retina, pero el proceso químico en cada uno que apoya la fototransducción es similar.[1] Durante la década de 1990 se descubrió una tercera clase de células fotorreceptoras:[2] el células fotosensibles del ganglio. Estas células no contribuyen a la vista directamente, pero se cree que la ayuda ritmos circadianos y reflejo pupilar.
Existen grandes diferencias funcionales entre los bastones y conos. Barras son extremadamente sensibles y pueden ser desencadenadas por un fotón.[3][4] En niveles muy bajos de luz, experiencia visual se basa únicamente en la señal de barra. Esto explica por qué no se pueden ver colores en bajos niveles de luz: sólo un tipo de célula del fotorreceptor es activo.
Conos requieren luz significativamente más brillante (es decir, un mayor número de fotones) para producir una señal. En los seres humanos, hay tres tipos diferentes de células cono, distinguido por su patrón de respuesta a diferentes longitudes de onda de la luz. Experiencia de color se calcula a partir de estas tres señales distintas, quizás a través de un proceso oponente.[5] Los tres tipos de células cono (áspero) responden a la luz de corto, mediano y largos longitudes de onda. Tenga en cuenta que, debido a la principio de univariance, la leña de la célula depende de la cantidad de fotones absorbidos. Las diferentes respuestas de los tres tipos de células cono se determinan por las probabilidades que sus proteínas fotorreceptoras respectivos absorben fotones de diferentes longitudes de onda. Así, por ejemplo, una célula del cono L contiene una proteína del fotorreceptor que más fácilmente absorbe longitudes de onda largas de luz (es decir, más "rojo"). Luz de longitud de onda más corta también puede producir la misma respuesta, pero debe ser mucho más brillante para hacerlo.
La retina humana contiene aproximadamente 120 millones barra células y 6 millones células del cono. El número y proporción de barras conos varía entre especies, depende de si un animal es principalmente diurnal o nocturnal. Algunos búhos, tales como la cárabo común,[6] tienen un enorme número de barras en sus retinas. Además, hay unos 2,4 millones células del ganglio en el sistema visual humano, los axones de estas células forman el 2 nervios ópticos, 1 a 2% de ellos fotosensible.
Las glándulas pineales y parapineal son fotorreceptoras en los vertebrados no mamíferos, pero no en mamíferos. Las aves tienen fotoactivos del líquido cerebroespinal (CFS)-ponerse en contacto con las neuronas dentro del órgano paraventricular responden a la luz en ausencia de entrada de los ojos o neurotransmisores. Fotorreceptores invertebrados en organismos tales como insectos y moluscos son diferentes en su organización morfológica y sus vías bioquímicas subyacentes. Se describen aquí son humano fotorreceptores.
Contenido
- 1 Histología
- 2 Seres humanos
- 3 Fototransducción
- 3.1 Corriente oscura
- 3.2 Vía de transducción de señal
- 3.3 Ventajas
- 3.4 Diferencia entre bastones y conos
- 4 Función
- 5 Desarrollo
- 6 Señalización
- 7 Fotorreceptores (no-no conos) de la célula del ganglio
- 8 Véase también
- 9 Referencias
- 10 Bibliografía
- 11 Enlaces externos
Histología
![Anatomy of a Rod Cell[7]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bb/Rod%26Cone.jpg/200px-Rod%26Cone.jpg)
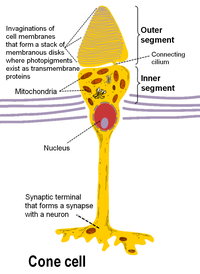
Barra de y cono fotorreceptores se encuentran en la capa más externa de la retina; ambos tienen la misma estructura básica. Más cercano al campo de visión (y más alejado el cerebro) es la Axon terminal, que libera una neurotransmisor llama glutamato Para células bipolares. Más atrás está el cuerpo de la célula, que contiene la celda organelos. Parte posteriora más lejana todavía es el segmento interno, una parte especializada de la célula lleno de mitocondrias. La función principal del segmento interno es proporcionar ATP (energía) para la bomba sodio-potasio. Por último, más cercana a la cerebro (y más alejado del campo de visión) es el segmento externo, la parte de los fotorreceptores que absorbe luz. Segmentos exteriores son realmente modificados cilios[8][9] contienen discos llenados de opsina, la molécula que absorbe fotones, así como voltaje-bloqueado canales de sodio.
La membranosa proteína del fotorreceptor opsina contiene una molécula de pigmento llamada retiniana. En las células de la barra, estas juntas se llaman rodopsina. En las células del cono, hay diferentes tipos de opsins que combinan con la retina en forma de pigmentos llamados photopsins. Tres clases diferentes de photopsins en los conos reaccionan a diversas gamas de frecuencia de luz, una diferenciación que permite que el sistema visual calcular el color. La función de la célula del fotorreceptor es convertir la energía luminosa de los fotones en una forma de energía fácilmente utilizable para el organismo y transmisibles del sistema nervioso: esta conversión se llama transducción de la señal.
El opsin encontró en las células fotosensibles del ganglio de la retina que intervienen en varias respuestas reflexivas del cerebro y del cuerpo a la presencia de la luz (día), como la regulación de la ritmos circadianos, reflejo pupilar y otras respuestas no visual a la luz, se llama melanopsina. Anormal en los vertebrados, melanopsina funcionalmente se asemeja a opsins invertebrados. En la estructura, es una opsina, un proteína de retinylidene gran variedad de Receptor g-proteína-juntado.
Cuando la luz activa la melanopsina sistema de señalización, el ganglio que contienen melanopsina células de descarga impulsos nerviosos que se llevan a cabo a través de sus axones metas específicas del cerebro. Estos objetivos incluyen la núcleo pretectal olivary (un centro responsable de control de la pupila del ojo), el LGN y, a través del zona retinohypothalamic (RHT), el núcleo supraquiasmático del hipotálamo (el marcapasos maestro de los ritmos circadianos). Que contienen melanopsina ganglio las células se cree que influyen en estos objetivos soltando sus terminales del axon de la neurotransmisores glutamato y pituitaria adenilato ciclasa, activación de polipéptido (PACAP).
Seres humanos



El ser humano retina tiene aproximadamente 6 millones conos y 120 millones de bastones.[12] Las señales de los bastones y conos convergen en el ganglio y las células bipolares para el preprocesamiento antes de ser enviados a la núcleo geniculado lateral. En el "centro" de la retina (el punto directamente detrás de la lente) se encuentra el fóvea (o fovea centralis), que contiene sólo células del cono; y es capaz de producir el más alto la región agudeza visual o más alto resolución. En el resto de la retina, bastones y conos se entremezclan. No hay fotorreceptores se encuentran en el punto ciego, el área donde las fibras de células del ganglio se recogen en el nervio óptico y dejan el ojo.[13]
Las proteínas del fotorreceptor en los tres tipos de conos difieren en su sensibilidad a los fotones de diferentes longitudes de onda (ver gráfico). Puesto que los conos responden a la longitud de onda y la intensidad de la luz, sensibilidad del cono a la longitud de onda se mide en términos de su tasa relativa de respuesta si la intensidad de un estímulo se mantiene fija, mientras que la longitud de onda es muy variada. De esto, a su vez, se infiere la absorbancia.[14] El gráfico normaliza el grado de absorbencia en escala de cien puntos. Por ejemplo, la respuesta relativa del cono S picos alrededor de 420 nm (nanómetros, una medida de longitud de onda). Esto nos indica que un cono de S es más probable que absorba un fotón en 420 nm que cualquier otra longitud de onda. Si la luz de diferente longitud de onda a la que es menos sensible, dice 480 nm, se aumenta en brillo adecuadamente, sin embargo, producirá exactamente la misma respuesta en el cono S. Por lo tanto, los colores de las curvas son engañosos. Conos no pueden detectar el color por sí mismos; por el contrario, visión de color requiere la comparación de la señal en los tipos de cono diferentes.
Fototransducción
El proceso de fototransducción se produce en el retina.[12] El retina tiene muchas capas de diversos tipos celulares.[12] Los más conocidos fotorreceptor células de (barras de y conos de) forman la capa más externa, más cercana a la escleray furthest desde el pupila.[12] Son los fotorreceptores responsables de vista. La capa media contiene células bipolares, que recogen las señales neurales de la barras de y de la conos de y luego transmitirlos a la capa más interna de la retina,[12] donde la neuronas llama retiniana ganglio de células de (RGCs), un pequeño porcentaje de los cuales ellos mismos son fotosensibles, organizar las señales y enviarlas al cerebro.[12] La liada RGC axones forma de la del nervio óptico, que sale del ojo a través de un agujero en el retina creación de la punto ciego.[12]
Activación de bastones y conos es realmente hiperpolarización; cuando no están siendo estimulados, ellos despolarizar y soltar continuamente glutamato. En la oscuridad, las células tienen una concentración relativamente alta de 3' - 5' monofosfato de guanosina cíclico (cGMP), que se abre canales del ion (en gran parte canales de sodio, aunque calcio puede entrar a través de estos canales también). Las cargas positivas de la iones que entrar en la célula por su Gradiente electroquímico cambio de la célula potencial de membrana, causar despolarización y conducir a la liberación del neurotransmisor glutamato. El glutamato puede despolarizar algunas neuronas y hyperpolarize otras.
Cuando la luz golpea un pigmento fotorreceptoras dentro de la célula del fotorreceptor, el pigmento cambia de forma. El pigmento, llamado iodopsin o rodopsina, consta de grandes proteínas llamadas opsin (situado en la membrana del plasma), unido a un grupo prostético covalentemente: una molécula orgánica llamó retiniano (un derivado de la vitamina A). La retina existe en la forma 11-cis-retinal cuando en la oscuridad, y la estimulación por la luz hace su estructura cambiar a todo-trans-retinal. Este cambio estructural hace que se active una proteína reguladora llamada transducina, que conduce a la activación de fosfodiesterasa de GMPC, que rompe el cGMP en 5'-GMP. Reducción de cGMP permite los canales del ion cerrar, prevenir la afluencia de iones positivos, hyperpolarizing la célula y detener la liberación de neurotransmisores.[15] Todo el proceso por que luz inicia una respuesta sensorial se llama fototransducción visual.
Corriente oscura
Sin estimular (en la oscuridad), nucleótido cíclico cerrados canales en el segmento externo están abiertos porque GMP cíclico (cGMP) está ligada a ellos. Por lo tanto, iones con carga positiva (es decir sodio iones) entre el fotorreceptor, despolarizantes que acerca de −40 milivoltio (potencial de reclinación en otras células del nervio está generalmente −65 mV). Esta despolarización actual a menudo se conoce como corriente oscura.
Vía de transducción de señal

El transducción de la señal es el mecanismo por el cual la energía de un fotón señala un mecanismo en la célula que conduce a su polarización eléctrica. Esta polarización conduce finalmente a la transmisión o inhibición de la señal neural que será alimentada al cerebro a través de la del nervio óptico. Los pasos o la vía de transducción de señal, en la barra y el cono del ojo vertebrado fotorreceptores son entonces:
- El rodopsina o iodopsin en el disco de membrana del segmento externo absorbe un fotón, cambia la configuración de un retiniana Base de Schiff cofactor dentro de la proteína de la forma cis a la trans-forma, causando la retina cambiar de forma.
- Esto se traduce en una serie de productos intermedios inestables, el último de los cuales se une más fuerte para un Proteína G En membrana, llamado transducinay lo activa. Este es el primer paso de amplificación – cada fotoactivado rodopsina activa activación de cerca de 100 transducins.
- Entonces cada transducina activa la enzima cGMP-específica fosfodiesterasa (PDE).
- PDE entonces cataliza la hidrólisis de GMPC a 5' GMP. Este es el segundo paso de amplificación, donde un solo PDE hidroliza alrededor de 1000 moléculas de cGMP.
- La neta concentración de GMPC intracelular es reducida (debido a su conversión a 5' GMP a través de PDE), resultando en el cierre de Na nucleotide-bloqueado cíclico+ canales iónicos en la membrana del segmento externo de fotorreceptor.
- Como resultado, los iones de sodio ya no pueden entrar en la célula y la membrana del segmento externo de fotorreceptor se convierte hyperpolarized, debido a la carga dentro de la membrana cada vez más negativa.
- Este cambio en el potencial de membrana de la célula hace que los canales de calcio voltaje-bloqueado cerrar. Esto conduce a una disminución en la afluencia de los iones del calcio en la célula y así baja la concentración de iones de calcio intracelular.
- Una disminución en la concentración de calcio intracelular significa que menos glutamato es liberado mediante exocitosis inducida por el calcio a la célula bipolar (véase abajo). (El nivel de disminución del calcio retarda la liberación del neurotransmisor glutamato, que excita la postsináptica células bipolares y células horizontales)
- Reducción en la liberación de glutamato significa una población de células bipolares será depolarized y una población separada de células bipolares será ser hyperpolarized, dependiendo de la naturaleza de los receptores (ionotrópicos o metabotrópicos) en la terminal postsináptica (véase campo receptivo).
Así, una varilla o cono de fotorreceptor realmente libera menos neurotransmisor al ser estimulados por la luz. Tampoco podría estimular menos neurotransmisor (despolarizar) o inhibir (hyperpolarize) los bipolares células sinapsis con, depende de la naturaleza del receptor en la célula bipolar. Esta capacidad es esencial para el centro de la asignación de las unidades visuales de encendido/apagado.[citación necesitada]
ATP, proporcionado por el segmento interno alimenta la bomba sodio-potasio. Esta bomba es necesaria para restablecer el estado inicial del segmento externo tomando los iones de sodio entran en la célula y les bombeo vuelve a salir.
Aunque los fotorreceptores son neuronas, que no llevan a cabo potenciales de acción con la excepción de la célula fotosensible del ganglio – que están implicadas principalmente en la regulación de la ritmos circadianos, la melatonina y la dilatación de la pupila.
Ventajas
Fototransducción en bastones y conos es único en que el estímulo (en este caso, claro) realmente reduce tasa respuesta o leña de la célula, que es inusual para un sistema sensorial donde el estímulo generalmente aumenta la tasa de respuesta o leña de la célula. Sin embargo, este sistema ofrece varias ventajas dominantes.
En primer lugar, el fotorreceptor (barra o cono) clásico es despolarizado en la oscuridad, lo que significa que fluyen muchos iones de sodio en la célula. Así, la apertura al azar o el cierre de canales de sodio no afecta el potencial de membrana de la célula; sólo el cierre de un gran número de canales, a través de la absorción de un fotón, se afectan y que la luz es en el campo visual de la señal. Por lo tanto, el sistema es silencioso.
En segundo lugar, hay una amplificación en dos etapas de fototransducción clásico: uno pigmento activar muchas moléculas de transducina, y un PDE se muchos cGMP. Esta amplificación significa que incluso la absorción de un fotón afectará el potencial de la membrana y la señal al cerebro que la luz es en el campo visual. Esta es la característica principal que distingue fotorreceptores de varilla de fotorreceptores del cono. Las barras son extremadamente sensibles y tienen la capacidad de registrar un solo fotón de luz, a diferencia de los conos. Por otra parte, los conos son conocidos por tener cinética muy rápida en términos de tasa de amplificación de fototransducción, a diferencia de las barras.
Diferencia entre bastones y conos
Comparación de la barra humana y células de cono, de Eric Kandel et en Principios de la ciencia Neural.[15]
| Barras de | Conos de |
|---|---|
| Utilizado para visión escotópica (visión bajo condiciones de poca luz) | Utilizado para visión fotópica (visión bajo condiciones de luz altas) |
| Muy ligero sensible; sensible a la luz dispersada | No muy fotosensible; sensible a la luz directa sólo |
| Causas de pérdida ceguera de noche | Causas de pérdida ceguera legal |
| Baja agudeza visual | Alta agudeza visual; mejor resolución espacial |
| No presentes en fóvea | Se concentró en fóvea |
| Respuesta lenta a la luz, añadidos con el tiempo los estímulos | Rápida respuesta a la luz, puede percibir cambios más rápidos en estímulos |
| Tienen más pigmento que conos, por lo que puede detectar niveles de luz más bajos | Tienen menos pigmento que las barras, requieren más luz para detectar imágenes |
| Pilas de discos de membrana-incluido son unattached a la membrana de la célula directamente | Los discos se unen a la membrana externa |
| Cerca de 120 millones barras distribuidas alrededor de la retina[12] | Unos 6 millones conos distribuidos en cada retina[12] |
| Un tipo de fotosensible pigmento | Tres tipos de fotosensible pigmento en los seres humanos |
| Confiere la visión acromática | Confiere la visión de color |
Función
No señal de fotorreceptores Color; sólo señalan la presencia de luz en el campo visual.
Un fotorreceptor dado responde a ambos la longitud de onda y intensidad de una fuente de luz. Por ejemplo, la luz roja con una cierta intensidad pueden producir la misma respuesta exacta en un fotorreceptor como luz verde de diferente intensidad. Por lo tanto, la respuesta de un solo fotorreceptor es ambigua cuando se trata de color.
Para determinar el color, el sistema visual compara las respuestas a través de una población de fotorreceptores (específicamente, los tres diferentes conos con diferentes espectros de absorción). Para determinar la intensidad, el sistema visual calcula cuántos fotorreceptores están respondiendo. Este es el mecanismo que permite visión de color tricromática en los seres humanos y algunos otros animales.
Desarrollo
Los eventos clave mediación rod versus cono S versus diferenciación cono M son inducidos por varios factores de transcripción, incluyendo RORbeta, OTX2, NRL, CRX, NR2E3 y TRbeta2. La suerte de cono S representa la programación predeterminada de fotorreceptor, sin embargo diferencial actividad transcripcional puede traer sobre barra o M cono generación. Conos de L están presentes en los primates, sin embargo es poco conocido por su programa de desarrollo debido al uso de roedores en la investigación. Hay cinco pasos para el desarrollo de fotorreceptores: proliferación de células progenitoras de retina multi-potentes (RPC); restricción de la competencia de RPC; Especificación de destino celular; expresión del gen fotorreceptor; y por último axonal crecimiento, formación de sinapsis y crecimiento del segmento externo.
Temprano Muesca señalización mantiene progenitor ciclismo. Precursores del fotorreceptor surge a través de la inhibición de la señalización de Notch y aumento de la actividad de varios factores incluyendo homólogo achaete-scute 1. OTX2 actividad compromete las células a la suerte del fotorreceptor. CRX más define el panel de fotorreceptores específicos de genes expresados. Expresión de NRL conduce a la suerte de vara. NR2E3 más restringe las células a la suerte de vara reprimiendo genes de cono. RORbeta es necesario para el desarrollo de la barra y el cono. TRbeta2 interviene la suerte de cono de M. Si cualquiera de las funciones de los factores anteriormente mencionados son la ablación, el fotorreceptor por defecto es un cono de S. Estos eventos tienen lugar en períodos de tiempo diferentes para especies diferentes e incluyen un patrón complejo de actividades que llevan a un espectro de fenotipos. Si se interrumpen estas redes de regulación, pigmentosa de la retinitis, degeneración macular u otros déficits visuales pueden resultar.[16]
Señalización
Los fotorreceptores de la barra y el cono de señal su absorción de fotones a través de una disminución en la liberación del neurotransmisor glutamato a células bipolares en su axon terminal. Puesto que el fotorreceptor es despolarizado en la oscuridad, se publica una gran cantidad de glutamato a células bipolares en la oscuridad. Absorción de un fotón hyperpolarize el fotorreceptor y por lo tanto el resultado en la liberación de menos glutamato en la presinápticos terminal de la célula bipolar.
Cada fotorreceptor de la barra o cono libera el mismo neurotransmisor, el glutamato. Sin embargo, difiere el efecto del glutamato en las células bipolares, dependiendo del tipo de receptor de incrustados en el membrana de la célula. Cuando el glutamato se une a un receptores ionotrópicos, la célula bipolar que despolarizar (y por lo tanto se hyperpolarize con luz como se lanza menos glutamato). Por otro lado, Unión de glutamato a un receptores metabotrópicos resultados en una hiperpolarización, por lo que esta célula bipolar a despolarizar a la luz como se lanza menos glutamato.
En esencia, esta propiedad permite una población de células bipolares que es excitada por la luz y otra población que es inhibido por él, aunque los fotorreceptores muestran la misma respuesta a la luz. Esta complejidad se hace importante y necesario para la detección de color, contraste, bordes, etc..
Mayor complejidad surge de las diferentes interconexiones entre células bipolares, células horizontales, y células amacrinas en la retina. El resultado final es diferentes poblaciones de células ganglionares en la retina, una subpoblación de los cuales es también intrínseco fotosensible, usando la melanopsina fotopigmento.
Fotorreceptores (no-no conos) de la célula del ganglio
Una varilla no no cono fotorreceptores en los ojos de los ratones, que fue demostrado para mediar ritmos circadianos, fue descubierto en 1991 por Foster et al.[2] Estas células neuronales, llamadas células ganglionares de la retina intrínsecamente fotosensibles (ipRGC), son un subconjunto pequeño (~ 1-3%) de la células ganglionares de la retina situado en el interior retina, es decir, en el frente[17] de las varillas y conos en la retina externa. Estas neuronas sensibles luz contienen un fotopigmento melanopsina,[18][19][20][21][22] que tiene un pico de absorción de la luz en diferentes longitudes de onda (~ 480 nm[23]) que los bastones y conos. Al lado de las funciones circadianas / comportamiento, ipRGCs tienen un papel en la iniciación de la reflejo pupilar de la luz.[24]
Dennis Dacey con colegas demostradas en una especie de mono de viejo mundo que las células gigantes del ganglio que expresan melanopsina proyectan el núcleo articulado lateral (LGN).[25] Previamente se habían demostradas solamente proyecciones al hipotálamo (núcleo supraquiasmático) y mesencéfalo (núcleo pre-tectal). Sin embargo un papel visual para el receptor fue todavía insospechado y no probado.
En 2007, Farhan H. Zaidi y los colegas publicaron trabajo pionero con los seres humanos coneless rodless. Current Biology posteriormente anunció en su editorial 2008, comentario y envíos a los científicos y los oftalmólogos, que el fotorreceptor no cono de barra no se había descubierto concluyente en seres humanos usando hito experimentos en seres humanos coneless rodless por Zaidi y los colegas[22][26][27][28] Como se comprobó en otros mamíferos, la identidad de la barra no no cono fotorreceptores en seres humanos fue encontrada para ser una célula del ganglio en la retina interna. Los trabajadores habían seguido por pacientes con enfermedades raras limpiando hacia fuera la función clásica de fotorreceptores de varilla y cono pero preservando la función de la célula del ganglio.[26][27][28] A pesar de no tener varillas o conos de los pacientes continúan exhiben photoentrainment circadiano, patrones de comportamiento circadianos, supresión de melanopsina y reacciones de la pupila, con sensibilidades espectrales pico a la luz ambiental y experimental que empareja para la melanopsina fotopigmento. Sus cerebros también podrían asociar visión con la luz de esta frecuencia.
En seres humanos el fotorreceptor de células ganglionares de la retina contribuye a consciente vista así en cuanto a funciones no forman imagen como ritmos circadianos, comportamiento y pupila reacciones.[29] Puesto que estas células responden sobre todo a la luz azul, se ha sugerido que tienen un papel en visión mesópica.[citación necesitada] Zaidi y trabajo de mis colegas con seres humanos coneless rodless por lo tanto, también abrieron la puerta en forma de imagen (visuales) papeles para el fotorreceptor de la célula del ganglio. Se descubrió que existen vías paralelas para la visión – una barra clásica y vía basada en el cono de la retina externa, y la otra una vía detector de luminosidad visual rudimentario derivadas de la retina interna, que parece ser activado por la luz antes que el otro.[29] Fotorreceptores clásicos también alimentan el sistema de la novela fotorreceptor, y constancia de color puede ser un papel importante como sugerido por Foster. El receptor podría ser instrumental en entender muchas enfermedades incluyendo la principales causas de ceguera en el mundo como el glaucoma, una enfermedad que afecta a las células del ganglio, y el estudio de los receptores ofrece potencial como una nueva vía para explorar en tratar de encontrar tratamientos para la ceguera. Es en estos descubrimientos del novela fotorreceptor en humanos y en el papel de los receptores en la visión, en lugar de sus funciones no forman imagen, donde el receptor puede tener mayor impacto en la sociedad como un todo, aunque el impacto de los ritmos circadianos alterados es otra área de relevancia para la medicina clínica.
Mayor parte del trabajo sugiere que la sensibilidad espectral máxima del receptor entre 482 y 460 nm. Steven Lockley et en 2003 demostró que 460 nm las longitudes de onda de la luz suprimen la melatonina dos veces tanto como más luz nm 555. Sin embargo, en trabajos más recientes por Farhan Zaidi et al., usando a seres humanos coneless rodless, fue encontrado que conscientemente lo que llevó a la percepción de la luz era un estímulo muy intenso de nm 481; Esto significa que el receptor, en términos visuales, permite una visión rudimentaria máximo para la luz azul.[29]
Véase también
- Fototransducción visual
- Receptor acoplado a proteínas G
- Sistema sensorial
- Fotosensible
- Célula fotosensible del ganglio
- Célula horizontal
- Célula bipolar
- Células amacrinas
Referencias
- ^ "ojo, humano." Encyclopædia Britannica. Encyclopaedia Britannica Ultimate Suite de referencia. Chicago: Encyclopædia Britannica, 2010.
- ^ a b Foster, R.G.; Provencio, I.; Hudson, D.; Fiske, S.; Agarre, w el.; Menaker, M. (1991). «Photoreception circadiano en el ratón retinally degenerado (rd/rd)». Diario de la fisiología comparativa A 169 (1): 39-50. doi:10.1007/BF00198171. PMID1941717.
- ^ Hecht, S.; Shlar, S.; Pirenne, M.H. (1942). "Energía, Quanta y visión". Diario de la fisiología General 25:: 819-840. doi:10.1085/JGP.25.6.819. PMC2142545. PMID19873316.
- ^ Baylor, D.A.; Cordero, T.D.; Yau, K.W. (1979). "Respuestas de la retina a fotones individuales". El diario de la fisiología 288:: 613-634. PMID112243.
- ^ Hurvich, Leo (1981). Visión de color. Sinauer.
- ^ "Vista del buho" en owls.org
- ^ Fisiología humana y los mecanismos de la enfermedad por Arthur C. Guyton (1992) ISBN 0-7216-3299-8 p. 373
- ^ Richardson, T.M. (1969). "Conexiones citoplásmicas y ciliares entre los segmentos internos y externos de los mamíferos receptores visuales". Investigación de visión 9:: 727-731. doi:10.1016/0042-6989 (69) 90010-8.
- ^ Louvi, A.; Arboleda, e. A. (2011). «Cilios en el SNC: el organelo tranquila reclama protagonismo ". Neurona 69:: 1046-1060. doi:10.1016/j.Neuron.2011.03.002.
- ^ Bowmaker J.K. y Dartnall H.J.A. (1980). "Pigmentos visuales de los bastones y conos en una retina humana". J. Physiol 298:: 501-511. PMC1279132. PMID7359434.
- ^ Fundamentos de la visión, Brian A. Wandell
- ^ a b c d e f g h i Schacter, Daniel L. (2011). Psicología segunda edición. 41 Madison Avenue, New York, NY 10010: digno de editores. págs. 136 – 137. ISBN978-1-4292-3719-2.
- ^ Goldstein, Bruce E. (2007). Sensación y percepción (7 ed.). Thomson y Wadswoth.
- ^ Wandell, Brian A. (1995). Fundamentos de la visión. Sunderland, MA: Sinauer.
- ^ a b Kandel, e. R.; Schwartz, J.H.; Jessell, T.M. (2000). Principios de la ciencia Neural (4ª Ed.). Nueva York: McGraw-Hill. PP. 507-513. ISBN0-8385-7701-6.
- ^ Swaroop, Anand; Douglas Kim y Douglas Forrest (agosto de 2010). "Regulación transcripcional de fotorreceptor desarrollo y Homeostasis en la Retina mamífera". Comentarios de Nature Neuroscience 11:: 563 – 576. doi:10.1038/nrn2880.
- ^ Ver retina para obtener información sobre la estructura de la capa retiniana.
- ^ Provencio, I. et al., (2000-01-15). "Un opsin humano en la retina interna". El diario de Neurociencia 20 (2): 600-605. PMID10632589.
- ^ Hattar, S.; Liao, HW; Takao, M; Berson, DM; Yau, KW (2002). "Las células ganglionares de la retina que contiene melanopsina: arquitectura, proyecciones y fotosensibilidad intrínseca". Ciencia 295 (5557): 1065-70. Bibcode:2002Sci... H 295,1065. doi:10.1126/Science.1069609. PMC2885915. PMID11834834.
- ^ Melyan, Z.; Tarttelin, e. E.; Bellingham, J.; Lucas, r. J.; Hankins, M. W. (2005). "Además de melanopsina humano hace photoresponsive de las células mamíferas". Naturaleza 433 (7027): 741 – 5. Bibcode:2005Natur.433... 741M. doi:10.1038/nature03344. PMID15674244.
- ^ Qiu, Xudong; Kumbalasiri, Tida; Carlson, Stephanie M.; Wong, Kwoon Y.; Krishna, Vanitha; Provencio, Ignacio; Berson, David M. (2005). "Inducción de la fotosensibilidad por expresión heteróloga de melanopsina". Naturaleza 433 (7027): 745-9. Bibcode:2005Natur.433... 745Q. doi:10.1038/nature03345. PMID15674243.
- ^ a b Vangelder, R (2008). «Photoreception no Visual: percepción de luz sin vista ". Current Biology 18 (1): R38. doi:10.1016/j.Cub.2007.11.027. PMID18177714.
- ^ Berson, David M. (2007). "Fototransducción en fotorreceptores de la célula del ganglio". Pflügers Archiv - diario Europeo de la fisiología 454 (5): 849 – 55. doi:10.1007/s00424-007-0242-2. PMID17351786.
- ^ Lucas, Robert J.; Douglas, Ronald H.; Foster, G. Russell (2001). "Caracterización de un photopigment ocular capaz de controlar la constricción pupilar en ratones". Nature Neuroscience 4 (6): 621 – 6. doi:10.1038/88443. PMID11369943.
- ^ Dacey, Dennis M.; Liao, Hsi-Wen; Peterson, Beth B.; Robinson, R. Farrell; Smith, C. Vivianne; Pokorny, Joel; Yau, King-Wai; Gamlin, Paul D. (2005). "Expresan melanopsina células ganglionares en primates retina señal color e irradiancia y proyecto para el GNL". Naturaleza 433 (7027): 749-54. Bibcode:2005Natur.433... D 749. doi:10.1038/nature03387. PMID15716953.
- ^ a b Genova, Cathleen, Los seres humanos ocultos que carecen de bastones y conos conservan respuestas normales a los efectos no visuales de la luz. Cell Press, 13 de diciembre de 2007.
- ^ a b A. Coghlan Personas ciegas 'ver' amanecer y el atardecer. Nuevo científico, 26 de diciembre de 2007, edición 2635.
- ^ a b Noticias médicas hoy. Respuestas normales a los efectos no visuales de luz retenida por los seres humanos ocultos que carecen de bastones y conos. 14 de diciembre de 2007.
- ^ a b c Zaidi FH et al (2007). "Onda corta sensibilidad a la luz de la conciencia circadiana, pupilar y visual en los seres humanos que carecen de una retina externa.". Biología actual: CB 17 (24): 2122 – 8. doi:10.1016/j.Cub.2007.11.034. PMC2151130. PMID18082405.
Bibliografía
- Campbell, Neil A. y Reece, Jane B. (2002). Biología. San Francisco: Benjamin Cummings. PP. 1064-1067. ISBN0-8053-6624-5.
- Freeman, Scott (2002). Ciencias biológicas (2ª edición). Acantilados de Englewood, NJ: Prentice Hall. págs. 835-837. ISBN0-13-140941-7.
Enlaces externos
- Búsqueda NIF – célula del fotorreceptor a través de la Marco de información de Neurociencia
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Otras Páginas
- Amigo a amigo
- Alboroto de Crown Heights (categoria judios y judaismo en la ciudad de Nueva York)
- Reconocimiento de iris
- Salud belleza vida
- Fatiga (medico)
- Henry E. Hardtner
- 2013 en television americana
- Campo de vision
- Dennis Rodman (categoria baloncesto expatriados estadounidenses en Finlandia)
- Anarrhotus
- ReplayGain
- Dormitorios de mascotas
- Educacion en Second Life
- Prueba de capacidad de trabajo
- Lista de las misiones diplomaticas de Polonia
- Arroyos (album de 1999)